Thomas Francis Jr. coined the phrase “original antigenic sin” in 1960 to describe a phenomenon he had first observed in the 1940s, and the theological register was not incidental: his father was a Welsh steelworker who had studied for the ministry as a young man in South Wales before emigrating to the industrial towns of western Pennsylvania, and the son’s choice of “sin” over any number of available immunological descriptors reflected a conviction that what he was naming had a structure the existing vocabulary of antibody kinetics could not adequately represent. The phenomenon itself was specific: college students vaccinated against the 1946 influenza virus who were subsequently infected by the antigenically drifted 1947 variant produced antibodies not against the infecting strain but against older strains they had previously encountered, as though the immune system, confronted with a novel antigen, preferentially recalled its primary response rather than generating a de novo one. Francis was arguing that the immune system carries the mark of its first antigenic encounter the way Christian anthropology holds that the soul carries the mark of its first transgression: indelibly, structurally, in a way that conditions every subsequent adaptive response without being accessible to conscious correction.
What interests me about Francis’s coinage is not the metaphor itself but the structural claim embedded within it, one that contemporary immunology has both deepened and complicated. The phrase endured for six decades, though it obscured as much as it clarified. “Sin” implies error, a falling-short, and the long history of the concept was accordingly preoccupied with pathology: the immune system was doing something wrong by recalling outdated antibodies when it should have been generating new ones. Recent work has fundamentally reframed the phenomenon. What Francis observed was not a defect but the surface expression of a permanent architectural reorganization of the germinal center response, and the emerging consensus is that the term “immune imprinting” better captures both the mechanism and its ambivalent valence. The imprint established by the first viral encounter is not a mistake the immune system makes; it is a constraint the immune system becomes.
I want to take this reframing seriously, ie. taking the biology seriously before making any claims about theology. The analogy I am pursuing is not a loose metaphor between immune memory and cognitive stubbornness, the kind of thing one encounters in popular science writing where “the body remembers” is made to do vaguely poetic work. The analogy is structural and specific: the germinal center, the lymph node microstructure where B-cells undergo somatic hypermutation and affinity-based selection, is physically reorganized by the first antigenic encounter in ways that constrain all subsequent responses, and I am arguing that something formally analogous happens to the cognitive apparatus one brings to theological reasoning when that apparatus is first organized by a particular tradition. The person raised in a Jaʿfarī household who later studies Sunnī jurisprudence does not encounter Sunnī jurisprudence de novo; their theological germinal centers are occupied by memory cells generated during the primary Jaʿfarī encounter, and these memory cells competitively exclude the naïve engagement that would be necessary to apprehend the Sunnī framework on its own terms. The response that is generated is always a cross-reactive response, shaped by the first encounter’s architecture, always a modification of the original antibody rather than a new one.
The claim is harder than it might initially appear, because it depends on specifics of germinal center architecture that most accounts of immune memory do not emphasize.
I. The Germinal Center as Finite Architecture
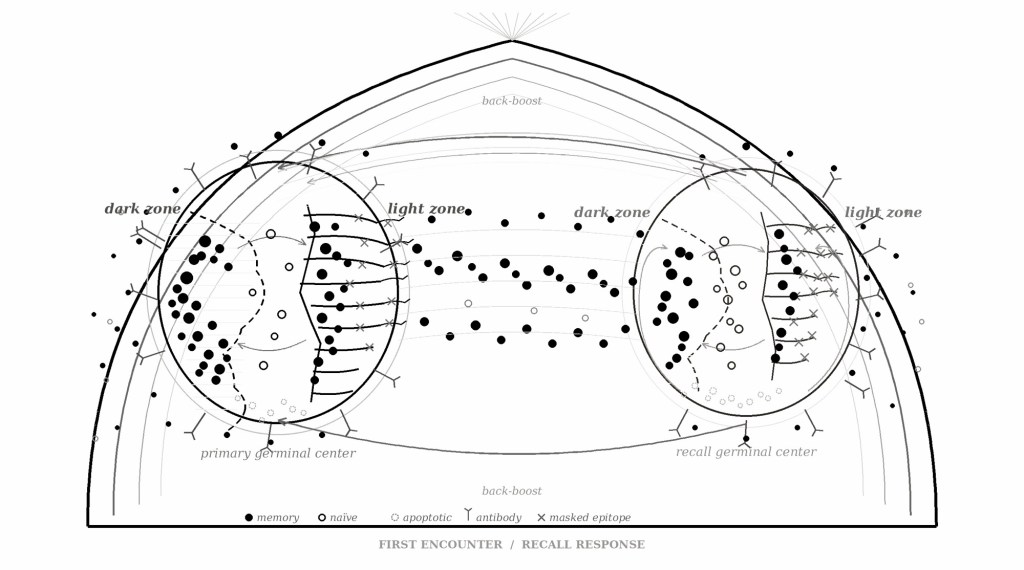
The germinal center is a transient microanatomical structure that forms within B-cell follicles of secondary lymphoid organs in response to T-dependent antigenic stimulation. It is where the adaptive immune system does its most sophisticated work: clonal expansion, somatic hypermutation, affinity-based selection, and differentiation into the memory B-cells and long-lived plasma cells that constitute durable humoral immunity. The structure is polarized into a dark zone, where centroblasts proliferate and activation-induced cytidine deaminase introduces point mutations into immunoglobulin variable-region genes at a rate roughly a million-fold above background, and a light zone, where centrocytes compete for antigen retained on follicular dendritic cell (FDC) surfaces and for limiting T-cell help. The entire system is a competitive ecology operating within fixed spatial boundaries, and like all competitive ecologies, it is shaped decisively by the order of arrival.
The insight that makes the analogy work at a structural level comes from Avancena et al., who demonstrated in a 2021 PNAS paper that mouse spleens contain a fixed number of FDC clusters per histological cross-section: identical across genetic backgrounds, unchanged by immunization or inflammation, present even in germ-free mice that have never encountered a foreign antigen. The architecture is predetermined. Immunization increases the occupancy of these pre-existing clusters but does not create new ones. When prior immunizations had pre-occupied the available FDC clusters with ongoing germinal center reactions, subsequent immunization with a completely novel antigen produced significantly impaired B-cell responses. The finite number of structural niches available for germinal center reactions means that prior immune responses physically occupy positions that are then unavailable for responses to new antigens. The constraint is spatial before it is functional.
If germinal center capacity were unlimited, if the immune system could open new germinal centers in response to new antigens without regard to prior occupancy, then imprinting would be a matter of preference or tendency, the kind of soft bias that might be overcome with sufficient exposure. The finitude of the architecture makes it something harder: a zero-sum competition for physical space in which memory cells from the first encounter have a structural advantage that no amount of subsequent exposure fully neutralizes. B-cells cycle iteratively between mutation in the dark zone and selection in the light zone, a process that Gabriel Victora’s laboratory at Rockefeller has mapped at single-cell resolution using in situ photoactivation and intravital multiphoton microscopy. Only a fraction of B-cells arriving in the light zone are positively selected to re-enter the dark zone for further rounds of mutation; the remainder undergo apoptosis or exit the germinal center as early memory cells or plasmablasts. The selection bottleneck is governed by the limiting availability of T follicular helper cells, themselves a scarce resource, so that higher-affinity B-cells systematically outcompete lower-affinity ones for survival signaling.
II. From Sin to Imprinting: What Changed and Why It Matters
The terminological shift from “original antigenic sin” to “immune imprinting” reflects three substantive revisions in understanding, not a cosmetic rebranding.
The first is that the phenomenon is not inherently pathological. Gostic et al. showed in a landmark 2016 Science paper that childhood hemagglutinin imprinting confers roughly seventy-five percent protection against severe infection and eighty percent protection against death from novel influenza subtypes in the same phylogenetic group. Hemagglutinin subtypes cluster into two groups: Group 1, which includes H1, H2, and H5, and Group 2, which includes H3 and H7. A person whose first influenza infection involved a Group 1 virus carries lifelong, substantial protection against H5N1; a person imprinted by Group 2 carries corresponding protection against H7N9. The protection tracks birth year rather than age, meaning the same birth cohorts remain at elevated risk for the mismatched group over decades of epidemiological observation. The cost is that the same imprinting that protects against matched-group threats leaves the individual disproportionately vulnerable to mismatched-group threats: protection purchased at the price of a specific vulnerability, a trade-off whose structure theology has its own language for.
The second revision is mechanistic. Francis described OAS as a phenomenon of antibody titers, a serological observation. Contemporary immunology has traced it to specific cellular and architectural mechanisms within the germinal center. Schiepers et al. in Victora’s laboratory published a 2023 paper in Nature using a molecular fate-mapping system to quantify the contribution of primary-encounter B-cells to recall antibody responses. Upon sequential homologous boosting, the overwhelming majority of serum antibodies derived from B-cells that had participated in the primary germinal center response. They called this “primary addiction,” a term that captures the compulsive quality of the recall response with more precision than “sin.” When boosting employed an antigenically distant variant, however, the balance shifted: a substantial fraction of antibodies came from newly recruited B-cells with superior variant-neutralizing activity. Antigenic distance, it turns out, can partially overcome imprinting, a finding whose analogue in theological cognition is the observation that traditions sufficiently distant from one’s own are sometimes easier to engage on their own terms than traditions that are close relatives.
The third revision, advanced most forcefully by Koutsakos and Ellebedy in a 2023 Immunity perspective piece, is a clarification of scope. OAS referred specifically to the dominance of the first encounter. Immune imprinting, as a concept, encompasses the cumulative effects of all prior encounters on subsequent responses. The immune system does not merely privilege its first response; it layers subsequent responses on top of the first in a stratified hierarchy that Lessler et al. formalized as “antigenic seniority” in a 2012 study analyzing neutralization titers against H3N2 strains across individuals spanning ages seven to eighty-one in Guangdong Province. Titers peaked for strains circulating when individuals were approximately seven years old and declined smoothly with age at first exposure. The immune autobiography is a palimpsest in which the earliest layers remain the most legible.
III. Competitive Exclusion: The Mechanism of Foreclosure
The cellular mechanism by which imprinting constrains subsequent responses is more layered than the simple narrative of memory cells “crowding out” naïve ones, and the specificity of the mechanism matters for the theological analogy.
Upon re-exposure to a related antigen, two populations of B-cells are available to respond: memory B-cells generated by the first encounter and naïve B-cells with no prior antigen experience. One might expect that both populations compete for entry into recall germinal centers, with memory cells winning by virtue of their higher-affinity, pre-mutated B-cell receptors. The actual picture, established by Mesin et al. in a 2020 Cell paper, is more surprising. Memory B-cells largely bypass the germinal center entirely upon recall, instead rapidly differentiating into extrafollicular plasmablasts that flood the serum with antibodies. Recall germinal centers, by contrast, are seeded predominantly by naïve B-cells: the vast majority of cells in secondary germinal centers had no prior germinal center experience. The recall antibody response, then, is dominated by memory-derived plasmablasts while recall germinal centers attempt, in parallel, to generate new diversity from naïve precursors.
This division of labor initially seems to soften the imprinting story. If naïve B-cells dominate recall germinal centers, perhaps the system can generate genuinely new responses after all. The complication is that the naïve B-cells entering recall germinal centers face a profoundly distorted selective environment. Pre-existing antibodies, circulating in serum and retained on FDC surfaces, physically sequester conserved epitopes, preventing naïve B-cell receptors from engaging them. This is epitope masking, and its effect is to narrow the antigenic territory available for de novo exploration. Schiepers et al. showed in a 2024 Immunity paper that pre-existing antibodies and memory T-cell help exert opposing forces on recall germinal center composition: antibody feedback narrows the response by masking familiar epitopes, while memory Tfh cells potentially diversify it by lowering the threshold for germinal center entry. The net result depends on the balance between these forces, but in practice, antibody feedback tends to dominate, particularly when the new antigen shares significant epitope overlap with the original.
The competitive exclusion, then, operates at two levels simultaneously. At the level of the immediate antibody response, memory B-cells overwhelm naïve responses by sheer speed and volume, producing cross-reactive antibodies that dominate serum within days. At the level of the germinal center, pre-existing antibodies reshape the selective landscape to constrain what kinds of new specificities can emerge. The foreclosure is neither total nor simple: it is a layered, context-dependent suppression whose severity scales with the antigenic distance between the original and the new encounter, the quantity of pre-existing antibody, and the availability of memory T-cell help. Absolute exclusion would be immunologically catastrophic. Partial exclusion, however, is sufficient to ensure that the organism’s response to any new but related threat bears the indelible signature of its first encounter.
A further, more recent finding reinforces this architectural bias. Barbulescu et al. showed in a 2026 Immunity paper that antibodies produced by the ongoing germinal center reaction itself feed back to modulate interclonal competition within that same reaction, reducing the abundance of clones targeting epitopes already covered by circulating antibodies and steering germinal center output toward novel epitopes. The system is not merely biased by its history; it is continuously reshaped by the antibodies its own history has produced.
IV. The COVID-19 Pandemic as a Natural Experiment in Imprinting
The COVID-19 pandemic provided an unintended but nearly ideal test of these mechanisms. Billions of people were first immunized with vaccines encoding the ancestral Wuhan-Hu-1 spike protein in 2021, and then confronted, beginning in late 2021, with the antigenically distinct Omicron lineage. The question was whether the immune systems of vaccinated individuals would generate de novo responses optimized against Omicron or whether their responses would be dominated by recall of Wuhan-imprinted memory.
The evidence converged rapidly. Park et al. showed in a 2022 Science paper that vaccination with ancestral spike followed by Omicron exposure preferentially recalled cross-reactive antibodies targeting conserved epitopes rather than generating Omicron-specific de novo responses. Bivalent boosters containing both ancestral and Omicron spike sequences fared no better: including ancestral spike appeared to be actively counterproductive, a finding that public health authorities eventually acted on by dropping the ancestral component entirely. The most striking result came from Tortorici et al. in a 2024 Immunity paper: a monovalent XBB.1.5 booster, containing no ancestral sequence whatsoever, still elicited responses dominated by recall of pre-existing Wuhan-reactive memory B-cells. A vaccine containing no ancestral sequence could not prevent the recall of ancestral-specific memory. The imprint had become constitutive: no longer a bias in the response but the architecture from which all responses were constructed.
The structural parallel to theological cognition is not that theological imprinting is as rigid or as mechanistically determined as immunological imprinting. The parallel is more precise than that: it illuminates the persistence of first-encounter architecture even when the person has explicitly and deliberately sought to move beyond it. The XBB.1.5 booster was designed to update the immune system, to teach it to recognize the new variant, and it partially succeeded, but the response it generated remained overwhelmingly shaped by an antigen that was no longer present in the formulation. The person who undertakes the study of a new theological tradition with full sincerity and intellectual commitment is doing something analogous: attempting to update their theological cognition, partially succeeding, but generating a response whose deep structure was laid down before the project of study began.
The concept of antigenic seniority provides the epidemiological frame. The hierarchy is ordinal, with each successive encounter taking its place below the previous ones. Gostic et al. extended this to pandemic influenza, showing that birth-year-specific imprinting with Group 1 or Group 2 hemagglutinin subtypes determines vulnerability to H5N1 versus H7N9 decades later, tracking birth year rather than age across nineteen years of surveillance data.
The theological equivalent is something every person of faith recognizes without being able to articulate precisely: the formative religious experiences of childhood and early adolescence retain an authority, a felt primacy, that no subsequent encounter fully displaces. The prayer learned at seven, the theological commitment absorbed at twelve, the scriptural passage memorized at fourteen: these are not merely earlier in the sequence; they are structurally dominant. Later theological encounters are layered on top of them but do not reorganize the foundation. The immunological evidence suggests that this is not a failure of intellectual maturation or a sign that the person has not sufficiently interrogated their childhood commitments. It is the expected behavior of a system whose architecture was determined by its first encounter with the relevant class of antigens.
V. Back-Boosting, or the Theology of Reinforcement
Back-boosting is the phenomenon that maps most directly onto the theological experience I am trying to describe, and its mechanism is specific. It is the observation that re-exposure to a variant antigen boosts antibodies against the original strain rather than generating optimal antibodies against the variant. In influenza serology, this manifests as the persistent elevation of titers against childhood strains upon exposure to antigenically drifted adult strains. In COVID-19 immunology, it manifested dramatically: Aydillo et al. showed that SARS-CoV-2 infection boosted antibodies against conserved regions of seasonal betacoronaviruses, the common-cold coronaviruses most people encounter in childhood, and this back-boost negatively correlated with the induction of SARS-CoV-2-specific antibodies. The immune system, confronted with a novel pathogen, reached back to the coronaviruses of childhood and generated a response shaped by that earlier encounter.
The cross-reactive antibodies produced by back-boosting are often functional, sometimes protective, and occasionally superior to a de novo response because they target conserved epitopes that the pathogen cannot easily mutate away from. The Gostic et al. data on birth-cohort protection against pandemic influenza is perhaps the strongest evidence that imprinting-derived cross-reactivity can be lifesaving. The point is that the response is structurally incapable of being a de novo response. It is always a modification of the original, always constrained by the architecture laid down in the first encounter, and the degree to which it can be modified is itself bounded by the antigenic distance between the original and the new threat.
I dwell on back-boosting because it describes exactly what happens when a person formed within one theological tradition encounters another. I was raised in a Jaʿfarī household, which means that my primary theological encounter was with a particular constellation of commitments: the necessity of the Imāmate, the continuity of walāya as a structural principle of sacred history, the hermeneutical authority of the Ahl al-Bayt, the permissibility of taqiyya under duress, the theological coherence of mutʿa, the specifically Shīʿī theology of intercession. These commitments were not presented to me as one option among several; they constituted the entire available antigenic landscape during the period when my theological germinal centers were first activated. They organized the architecture of my religious cognition in the way that a first influenza infection organizes the B-cell repertoire: by generating memory cells whose specificity constrains all subsequent encounters.
When I later encountered Sunnī jurisprudence in an academic context, reading Shāfiʿī’s Risāla or working through Ghazālī’s al-Mustaṣfā, I did not encounter these texts as an unprimed reader would. I encountered them through the cross-reactive antibodies generated by my Jaʿfarī imprinting. Every Sunnī legal principle was processed by a cognitive apparatus already organized around the Jaʿfarī framework: the Sunnī theory of the caliphate was perceived through the lens of the Imāmate, the Sunnī hadith canon was evaluated against the Jaʿfarī criteria of transmission, the Sunnī position on consensus was read against the Jaʿfarī insistence on the continuing authority of the living Imām. My engagement was not hostile; I was not polemicizing. I was experiencing the theological equivalent of back-boosting: a new antigenic encounter that activated pre-existing memory cells and produced a response shaped overwhelmingly by the first encounter’s architecture.
Consider a concrete instance. The Sunnī doctrine of ijmāʿ, scholarly consensus, holds that the agreement of qualified jurists in a given generation constitutes a binding source of law, grounded in the hadith “my community will not agree upon an error.” For a person encountering this doctrine without prior theological commitment, the questions it raises are internal to Sunnī legal theory: what counts as consensus, who qualifies as a jurist, whether consensus once established can be overturned by subsequent generations. These are the questions the doctrine generates on its own terms. When I encounter the same doctrine, however, my cognitive apparatus does not generate these questions first. It generates a different set, ones legible as questions only within the architecture of my Jaʿfarī imprinting: if consensus is authoritative, what happens when it conflicts with the explicit ruling of an Imām? If the community cannot agree upon an error, does this apply only to the community that recognized the rightful succession, or to the community that deviated from it? The Sunnī jurist would not recognize these as the natural questions the doctrine raises; they are the cross-reactive antibodies produced by my memory cells, shaped to bind a different antigen but reaching for the nearest available epitope on this one. I can learn, over time, to ask the questions the doctrine generates on its own terms, and I can learn to set aside, temporarily and with effort, the questions my imprinting generates. This is affinity maturation. What I cannot do is encounter the doctrine as though the Jaʿfarī questions had never occurred to me, as though the memory cells were not present, as though the germinal centers of my theological cognition had not already been organized by a prior encounter with the question of authority in Islamic law.
The theological “antigen,” to be precise about a term I have been using loosely, is not a doctrine. It is not a propositional claim that could be evaluated in isolation the way one evaluates a syllogism. It is a hermeneutical orientation: a way of reading, a prior organization of the categories through which doctrines are received, before any particular doctrine is encountered. The Jaʿfarī antigen is not “the Imām is divinely appointed” as a proposition; it is the interpretive disposition that makes questions of authority, continuity, and esoteric meaning salient in every theological encounter, the disposition that determines which features of a new doctrinal landscape are recognized as epitopes and which remain invisible. This is why the analogy must be to the germinal center and not to a simpler model of memory: the immune system does not remember antigens the way a filing cabinet stores records. It is architecturally reorganized by them, so that the apparatus of recognition itself is different after the encounter than before. The theological antigen, likewise, is not a piece of information but a reorganization of the apparatus by which information is processed.
The phenomenon is symmetrical, which is part of what makes it inescapable. A Sunnī jurist encountering the Jaʿfarī theory of the Imāmate will process it through cross-reactive antibodies generated by their own imprinting: the Imāmate will be perceived as a theory of political authority (because the Sunnī framework already has a category for that), its theological dimensions will be assimilated to existing categories of sainthood or scholarly authority (because those are the nearest available epitopes), and its more distinctive features, the ones that have no cognate in the Sunnī framework, will be the most likely to be overlooked or flattened, because there are no pre-existing memory cells with even partial specificity for them. The symmetry matters because it means the constraint operates independently of the relative merits of the traditions in question. The issue is not which tradition is correct; it is that the cognitive apparatus for evaluating correctness has already been organized by the first tradition in ways that shape the evaluation itself.
The obvious objection, and the one the argument itself requires me to address while the autobiographical material is still proximate, is that the immunological framework through which I am reading theology is itself a cross-reactive response generated by my own cognitive imprinting. I trained as a physician before I wrote as an essayist, which means that the germinal center is not a neutral analogy I selected from among equally available options; it is the framework my own first-encounter architecture makes most available. A philosopher would reach for Gadamer’s Vorurteil; a sociologist for Bourdieu’s habitus; a cognitive scientist for predictive processing. Each of these captures some of the same structural features I have described, and each would emphasize different aspects of the phenomenon. The germinal center model is the one I find most adequate to the phenomenon, but “find most adequate” is itself a statement about the architecture of my cognitive apparatus rather than a claim about the objective superiority of the model. The immunological vocabulary feels precise to me because it was laid down in the germinal centers of my medical training, and the affinity of my cognitive antibodies for biological models is higher than their affinity for philosophical or sociological ones. I cannot fully correct for this, any more than the Jaʿfarī reader of Sunnī jurisprudence can fully correct for the questions their imprinting generates. What I can do is name it, which is what the argument itself demands.
VI. What Interfaith Dialogue Actually Is (Immunologically Speaking)
If the structural parallel holds, then the interfaith dialogue, the comparative jurisprudence seminar, the ecumenical theological project all proceed on an assumption the immunological evidence does not support: that mature minds can generate fresh, unbiased responses to unfamiliar theological frameworks, that with sufficient goodwill and intellectual discipline one can set aside one’s formative commitments and engage another tradition on its own terms.
What these enterprises actually accomplish is affinity maturation of cross-reactive antibodies.
By this I do not mean that the learning is trivial. In the germinal center, B-cells do not simply reproduce the antibodies generated in the primary response; they undergo further rounds of somatic hypermutation and affinity-based selection that gradually improve the fit between their first-encounter-derived antibodies and the new antigen. The antibodies become better, more capable of neutralizing the new threat, but they remain modifications of the original response rather than de novo constructions. When I spend sustained time with Sunnī legal theory, something happens: my understanding of it becomes more refined, more capable of tracking its internal logic, more sensitive to its distinctive emphases. I can, after sufficient engagement, reproduce a Sunnī argument for the authority of scholarly consensus in a way that a Sunnī interlocutor would recognize as accurate. This refinement is genuine learning. What it does not represent is a de novo response. The cognitive architecture generating my understanding of Sunnī consensus remains organized around the Jaʿfarī principle of Imāmic authority; my apprehension of the Sunnī position is always, at some level, a modification of my apprehension of the Jaʿfarī position, shaped by the question “how does this relate to what I already hold?” rather than the question “what does this mean on its own terms?”
One can observe the same dynamic in the academic study of religion. The Catholic scholar of Islam whose work is shaped, however subtly, by sacramental categories. The Protestant historian of Buddhism whose account of the Mahāyāna schools carries traces of Reformation historiography. The secular scholar whose treatment of mystical experience is organized around categories of explanation that a practitioner would not recognize as primary. These are sophisticated, refined, informative cross-reactive responses that remain structurally tethered to their point of origin.
VII. Antigenic Distance and the Paradox of the Far Tradition
The immunological research contains a finding that complicates this picture in a way that matters theologically. Schiepers et al. showed that antigenic distance modulates the severity of imprinting: when the new antigen is sufficiently different from the original, the balance shifts and a larger proportion of the response comes from genuinely new B-cell clones. Yisimayi et al. demonstrated that repeated exposures to Omicron variants can partially override ancestral SARS-CoV-2 imprinting, generating broader neutralization, though a single variant booster is insufficient.
The theological implication is counterintuitive. If the argument holds, then the traditions most difficult to engage on their own terms are the ones closest to one’s own: the Sunnī jurist’s encounter with Jaʿfarī jurisprudence, the Roman Catholic theologian’s encounter with Eastern Orthodoxy, the Reform Jew’s encounter with Conservative halakha. In each case, the antigenic overlap is enormous, cross-reactive memory cells will be maximally activated, and competitive exclusion of naïve engagement will be most severe. The response will be almost entirely composed of modified first-encounter antibodies, and the person will have the strongest subjective sense of understanding precisely because the cross-reactive response fits the new antigen reasonably well without being optimally targeted to it. Herein lies the peculiar danger of near-cognate traditions: the cross-reactivity is high enough to produce a convincing illusion of comprehension.
The uṣūlī concept of taʿāruḍ al-adilla, the conflict between equally weighted proofs, and the rules of tarjīḥ (preference) that resolve it, maps onto what happens when cross-reactive antibodies from two closely related traditions compete for the same epitope. In classical Sunnī uṣūl, tarjīḥ is presented as a neutral adjudicatory procedure: when two proofs of equal apparent weight conflict, the jurist applies a hierarchy of preference criteria to determine which prevails. The imprinting model suggests that tarjīḥ is never the neutral operation the uṣūlīs describe it as. The criteria of preference are themselves processed by a cognitive apparatus already organized by the jurist’s formative tradition, so that which proof appears “more specific” or “better attested” is shaped by the architecture of prior encounter. The Jaʿfarī jurist resolving a taʿāruḍ between a Sunnī proof and a Jaʿfarī proof does not adjudicate from a neutral vantage; the Jaʿfarī proof benefits from the higher affinity and faster activation kinetics of the memory cells that generated the categories through which “strength of attestation” is assessed. Tarjīḥ, in other words, is itself an imprinted operation, and this has consequences for every project of comparative jurisprudence that treats cross-school adjudication as procedurally neutral.
Traditions that are antigenically distant, by contrast, may be paradoxically more accessible to something approaching naïve engagement. The Jaʿfarī Muslim encountering Theravāda Buddhist soteriology, or the Lutheran theologian engaging with Hindu darśana, faces an antigenic landscape so different from their imprinting antigen that cross-reactive memory cells have little to bind. The response will be weaker and less confident, precisely because the first-encounter architecture provides less scaffolding, but it will contain a larger proportion of new cognitive specificities. The person may understand less initially but understand differently, with a smaller ratio of projection to perception. This maps onto the common pedagogical observation that introductory students sometimes ask questions about a foreign tradition that specialists, constrained by their own cross-reactive apparatus, would never think to ask.
The paradox has a cost. The enhanced engagement with distant traditions comes at the expense of reduced integration: precisely because the new response is less shaped by existing architecture, it is also less connected to the person’s broader cognitive immune system, less integrated into their network of commitments, more likely to remain an isolated pocket of understanding. This too has a biological correlate. Naïve B-cells that enter recall germinal centers targeting genuinely novel epitopes must compete for Tfh cell help against a background of cross-reactive clones, and the memory cells they eventually produce occupy a marginal position in the overall immune repertoire. They are real, they are functional, but they do not displace the dominant clones. A person’s encounter with a distant theology may produce real understanding without that understanding achieving integration into the deeper commitments that shape how they actually reason when the stakes are high.
VIII. The Conditions of Conversion (Immunologically Speaking)
If immune imprinting constrains all subsequent responses without absolutely foreclosing them, what would the immunological equivalent of a genuine theological conversion look like?
The first possible mechanism is massive, repeated exposure to the new antigen over extended periods. Yisimayi et al. showed that multiple Omicron exposures can partially override ancestral imprinting, and clinical data suggests that individuals with extensive variant exposure history develop broader neutralization profiles. The theological analogue would be deep, prolonged immersion in a second tradition: the years-long novitiate, the extended period of study under a master in a new tradition, the slow reshaping of daily practice and communal affiliation. Even here, the override is partial. The ancestral imprint is not erased but overlaid, and the resulting repertoire reflects the entire history of encounters in their chronological hierarchy. The convert does not become a person who was always of the new tradition; they become a person whose cognitive repertoire bears the layered imprint of both, with the first remaining structurally dominant even as the second occupies an increasing share of the active response.
The second mechanism is more radical. If the pre-existing antibodies that mediate epitope masking could be removed, naïve B-cells would have unimpeded access to the new antigen. There is no clean biological way to accomplish this in vivo, but the theoretical implication is that the removal of the mediating structures of the first tradition, the communities, practices, texts, and relationships that sustain the plausibility of the original framework, would be a precondition for something approaching naïve engagement with a new tradition. Peter Berger’s concept of “plausibility structures” maps onto this: the social networks that maintain the subjective plausibility of a belief system function like circulating antibodies that mask alternative epitopes, and their disruption, through migration, exile, social rupture, or deliberate withdrawal, creates the conditions under which a more de novo encounter becomes possible. The phenomenology of conversion frequently involves exactly such disruption: the convert does not simply add a new framework to an existing one but undergoes a period of cognitive immunosuppression in which the structures that sustained the first framework are weakened or dismantled. Al-Ghazālī’s account of his own crisis in the Munqidh min al-Ḍalāl reads, on this model, as a description of deliberate immunosuppression. The details matter: at the height of his career at the Niẓāmiyya in Baghdad, holding the most prestigious teaching post in the Islamic world, Ghazālī describes a period of roughly six months in which his tongue dried up, he could not eat, he could not lecture. The crisis was not propositional, not a matter of encountering a better argument; it was architectural, a collapse of the plausibility structures of Ashʿarī kalām and Aristotelian philosophy that had organized his theological cognition so thoroughly that their dissolution registered as a physical inability to function. He then abandoned his post and spent approximately eleven years in wandering and seclusion, in Damascus and Jerusalem and Mecca, before the conditions for a de novo encounter with Sufi experiential theology had been sufficiently prepared. The duration is itself significant: eleven years is not the timeline of a person evaluating arguments but of a person whose cognitive immune system required near-complete depletion before new germinal centers could form.
The immunological caution, however, is that systemic immunosuppression is dangerous precisely because it removes protective responses along with constraining ones. The person who dismantles their plausibility structures does not arrive at a neutral starting point; they arrive at a state of cognitive vulnerability in which the new tradition’s antigens encounter an apparatus stripped of its capacity for discrimination. This may account for the frequently observed intensity and rigidity of converts, whose engagement with their new tradition often has a quality of overcommitment that native adherents find puzzling: the de novo response, generated without the modulating influence of pre-existing cross-reactive antibodies, may paradoxically produce a narrower, less differentiated repertoire than the affinity-matured cross-reactive response of the lifelong practitioner. The immunological term would be something like a monoclonal response where a polyclonal one is called for: all the cognitive resources directed at a single epitope rather than distributed across the antigen’s full surface.
IX. Fiṭra and the Limits of the Analogy
The claim is structural, not literal. I am not arguing that theological cognition operates by somatic hypermutation and affinity-based selection. I am arguing that the germinal center and the cognitive apparatus for theological reasoning share a set of formal properties, finite capacity, competitive dynamics, architectural constraint by first encounter, path-dependent modification of subsequent responses, that make the germinal center a better model for theological cognition than the models we typically use.
The propositional model, which treats theology as though changing one’s commitments were a matter of evaluating arguments, fails because it cannot account for the architectural dimension. Arguments are universally accessible; the same syllogism is available to any reasoner regardless of their prior commitments. Theological cognition does not operate on arguments in this way. It operates on arguments as they are processed by a cognitive apparatus that has been organized by prior theological encounters. The same argument lands differently depending on the architecture that receives it, and this is not a deficiency to be corrected but a structural feature of how theological cognition works.
The social-psychological model, confirmation bias and motivated reasoning and identity-protective cognition, comes closer but still treats the phenomenon as a functional distortion, something the mind does to incoming information that could in principle be corrected by sufficient self-awareness. The immune imprinting model says something more radical: the constraint is in the architecture of the processing apparatus itself. You cannot overcome imprinting by trying harder, any more than the immune system can overcome primary addiction by mounting a more vigorous secondary response. The more vigorous the response, the more the existing memory cells dominate it. Effort amplifies the constraint it aims to overcome.
There is a theological tradition that understands this, though it uses different language. The Shīʿī concept of fiṭra, the primordial disposition, holds that human beings are born with an innate orientation toward divine reality that precedes and conditions all subsequent theological encounter. The concept has obvious parallels to the innate immune system, the set of pre-existing pattern-recognition receptors that shape the adaptive response before any specific antigen is encountered. But fiṭra, unlike the innate immune system, is held to be universally identical: all human beings share the same primordial disposition, and variation arises from the particular encounters that shape it. The hadith most commonly cited in this connection, reported in both Sunnī and Shīʿī collections, states that every child is born upon the fiṭra, and it is the parents who make the child a Jew, a Christian, or a Zoroastrian. The hadith is typically read as an assertion about the primacy of natural religion over acquired tradition, and the implication drawn from it is that beneath the layers of culturally specific religious formation there persists a universal, unconditioned orientation toward the divine that can, at least in principle, be recovered.
The immunological parallel complicates this reading. Neonatal passive immunity, the transfer of maternal IgG antibodies across the placenta during the third trimester, means that the infant’s immune system encounters its first antigenic landscape through a repertoire of antibodies it did not generate. These maternal antibodies shape the infant’s early immune responses by masking certain epitopes and leaving others available for de novo engagement, effectively determining which antigens the child’s own germinal centers will encounter first. The theological parallel is that the parents do not merely present the child with a tradition as the first antigen; they provide the antibody environment, the interpretive atmosphere of practice, language, assumption, and emotional association, within which the child’s own primary response occurs. The child does not encounter “Islam” or “Christianity” or “Judaism” as a naïve B-cell encounters a foreign antigen; it encounters the tradition through a pre-existing repertoire of parental antibodies that have already organized the antigenic landscape, masking certain features and exposing others. By the time the child’s own adaptive theological cognition activates, the environment in which it activates has already been shaped by a transferred repertoire. The shaping is one layer deeper than the hadith suggests.
The immune imprinting model raises a more troubling possibility: the shaping encounter is so thorough, so architecturally transformative, that the fiṭra effectively disappears beneath the imprint, becoming inaccessible as an independent source of theological cognition. One does not return to the primordial disposition by stripping away the layers of acquired theology; one discovers that the primordial disposition has been permanently reorganized by the first encounter and is no longer available in its unimprinted form. The hadith says the parents make the child a Jew, a Christian, or a Zoroastrian, but it does not say the child can be unmade; the immunological evidence suggests that unmaking would require the dismantling of the very architecture through which theological cognition operates, leaving the person not with a recovered fiṭra but with a depleted repertoire. The naïve B-cell pool after the germinal centers have completed their first reaction is not the same thing as the pre-immune B-cell pool: the cells that might have responded with genuine novelty have been outcompeted, driven to apoptosis, or differentiated into irrelevant lineages. What remains is not the original potential but a diminished version of it, constrained by the residue of the architecture that was.
There is a competing Shīʿī reading of fiṭra, developed most fully in the philosophical tradition of Mullā Ṣadrā and refined by ʿAllāma Ṭabāṭabāʾī in the al-Mīzān, which holds that fiṭra is not merely an initial state but an ongoing, dynamic orientation that can be strengthened or weakened but never entirely effaced. The Ṣadrian framework is more precise than the generic version of this claim: fiṭra on his account is not a static endowment but a mode of existential motion (ḥaraka jawhariyya) toward the Real, one that cannot be eliminated because it is constitutive of the soul’s substantial reality rather than an accidental property that could be removed. This is a more hopeful picture, and it may well be the correct one theologically. The immunological analogy does not support it cleanly: the innate immune system shapes the adaptive response but does not override it, and in the specific context of germinal center imprinting, innate signals modulate the intensity of the response without altering the fundamental competitive dynamics that determine which B-cell clones dominate it. The fiṭra, on this model, would be the background signal that conditions all theological cognition without determining which tradition’s memory cells occupy the germinal centers. It would be real, but it would not be sufficient to overcome imprinting.
This is, I think, the unsettling implication of the analogy, and I do not want to soften it. The interfaith dialogue, the comparative jurisprudence seminar, the ecumenical theological project: all of these are valuable, all of them produce genuine learning, and all of them operate within constraints that their participants rarely acknowledge. What they can do, and what they demonstrably achieve, is affinity-mature their cross-reactive antibodies, gradually improving the fit between their first-encounter-derived response and the new antigen. This is real, and it is important, and it is the best that can be achieved within the constraints of imprinting. The person who mistakes affinity-matured cross-reactivity for de novo understanding has mistaken a better-fitting antibody for a new one, and this mistake has consequences for how they evaluate their own comprehension of traditions other than their own.
I return to the language Francis chose. He called it a sin, reaching for the theological register to name something the immunological vocabulary of his time could not adequately express: the idea that a first encounter leaves a mark so deep that it conditions everything that follows, that the organism carries the consequences of an event it did not choose and cannot reverse, that the very architecture of response is altered in ways that preclude innocence. The immunologists who followed him have spent six decades clarifying that the mark is not a sin but an imprint, that it is as often protective as it is constraining, that it reflects the evolved optimization of a system operating under finite resources rather than a defect in design. These clarifications are correct, and they matter. But Francis’s instinct for the theological register was also correct, because the phenomenon he identified is one that theology has always understood better than immunology: the impossibility of a second innocence, the recognition that formation is irreversible, the knowledge that every encounter after the first is shaped by what the first encounter made of you.
Every theological encounter after the first is a recall response: faster than a primary response, shaped by prior architecture, dominated by memory cells generated in a different antigenic context. The learning that occurs within this constraint is genuine, substantial, and the primary means by which intellectual life proceeds. The humility the constraint demands is equally genuine: the recognition that one’s understanding of a tradition other than one’s own, however refined, remains a cross-reactive response whose architecture was laid down elsewhere, in the germinal centers of a childhood one did not choose and cannot undo.
Recommended Reading
Immunology: Primary Sources
- Avancena, P., Song, T., Yao, Y., et al. “The magnitude of germinal center reactions is restricted by a fixed number of preexisting niches.” PNAS 118, no. 30 (2021): e2100576118.
- Schiepers, A., van ’t Wout, M.F.L., Greaney, A.J., et al. “Molecular fate-mapping of serum antibody responses to repeat immunization.” Nature 615 (2023): 482–489.
- Schiepers, A., Van ’t Wout, M.F.L., Hobbs, A., Mesin, L., and Victora, G.D. “Opposing effects of pre-existing antibody and memory T cell help on the dynamics of recall germinal centers.” Immunity 57, no. 7 (2024): 1618–1628.
- Barbulescu, A., Bilanovic, J., Langelaar, T., et al. “Antibody-mediated feedback modulates interclonal competition in the germinal center.” Immunity 59, no. 3 (2026): 734–745.
- Mesin, L., Schiepers, A., Ersching, J., et al. “Restricted Clonality and Limited Germinal Center Reentry Characterize Memory B Cell Reactivation by Boosting.” Cell 180 (2020): 92–106.
- Gostic, K.M., Ambrose, M., Worobey, M., and Lloyd-Smith, J.O. “Potent protection against H5N1 and H7N9 influenza via childhood hemagglutinin imprinting.” Science 354 (2016): 722–726.
- Lessler, J., Riley, S., Read, J.M., et al. “Evidence for Antigenic Seniority in Influenza A (H3N2) Antibody Responses in Southern China.” PLoS Pathogens 8, no. 7 (2012): e1002802.
- Koutsakos, M. and Ellebedy, A.H. “Immunological imprinting: Understanding COVID-19.” Immunity 56, no. 5 (2023): 909–913.
Immunology: Reviews and Context
- Victora, G.D. and Nussenzweig, M.C. “Germinal Centers.” Annual Review of Immunology 40 (2022): 413–442.
- Henry, C., Palm, A.-K.E., Krammer, F., and Wilson, P.C. “From Original Antigenic Sin to the Universal Influenza Virus Vaccine.” Trends in Immunology 39, no. 1 (2018): 70–79.
- Francis, T. “On the Doctrine of Original Antigenic Sin.” Proceedings of the American Philosophical Society 104, no. 6 (1960): 572–578.
COVID-19 Imprinting
- Tortorici, M.A., Addetia, A., Seo, A.J., et al. “Persistent immune imprinting after XBB.1.5 COVID vaccination in humans.” Immunity 57, no. 4 (2024): 875–883.
- Park, Y.-J., Pinto, D., Walls, A.C., et al. “Imprinted antibody responses against SARS-CoV-2 Omicron sublineages.” Science 378 (2022): 619–627.
Theology and Philosophy of Religion
- Berger, P.L. The Sacred Canopy: Elements of a Sociological Theory of Religion. New York: Doubleday, 1967.
- Ṭabāṭabāʾī, M.Ḥ. al-Mīzān fī Tafsīr al-Qurʾān. Beirut: Muʾassasat al-Aʿlamī, various editions. [The discussion of fiṭra in the tafsīr of Sūrat al-Rūm 30:30 is the relevant section.]
- Mullā Ṣadrā (Ṣadr al-Dīn al-Shīrāzī). al-Asfār al-Arbaʿa. [The framework of substantial motion (ḥaraka jawhariyya) undergirds the dynamic reading of fiṭra as ongoing existential orientation.]
- al-Ghazālī. al-Munqidh min al-Ḍalāl (Deliverance from Error). [Readable as a first-person account of deliberate cognitive immunosuppression and subsequent re-imprinting.]
- Shāfiʿī, M. ibn Idrīs. al-Risāla. Trans. Majid Khadduri. Cambridge: Islamic Texts Society, 1961.
Interdisciplinary
- Gadamer, H.-G. Truth and Method. Trans. Joel Weinsheimer and Donald G. Marshall. London: Continuum, 2004. [The concept of Vorurteil (prejudgment) and the claim that understanding always proceeds from within a tradition is the philosophical position closest to the argument of this essay, though without the immunological specificity.]
- Kuhn, T.S. The Structure of Scientific Revolutions. Chicago: University of Chicago Press, 1962. [Kuhn allows for revolutionary replacement where the imprinting model suggests layering.]
- Polanyi, M. Personal Knowledge: Towards a Post-Critical Philosophy. Chicago: University of Chicago Press, 1958. [The concept of “tacit knowledge” provides a philosophical framework for the non-propositional dimensions of theological imprinting.]




Leave a comment